Las membranas biológicas
Estas definen los límites externos de las células, y separan compartimientos dentro de ellas. Son componentes esenciales de todas las células vivas. Una membrana típica está formada por dos capas de moléculas de lípidos y muchas proteínas embebidas en ella. Las membranas biológicas no sólo son barreras pasivas contra la difusión. Tienen una gran variedad de funciones complejas. Algunas proteínas contenidas en las membranas sirven como bombas selectivas que controlan en forma estricta el transporte de iones y de moléculas pequeñas que entran y salen de la célula. Las membranas también son responsables de generar y mantener la concentración de gradientes de protones, esenciales para la producción de ATP. Los receptores en las membranas reconocen señales extracelulares y las comunican al interior de la célula.
Bicapa lipídica
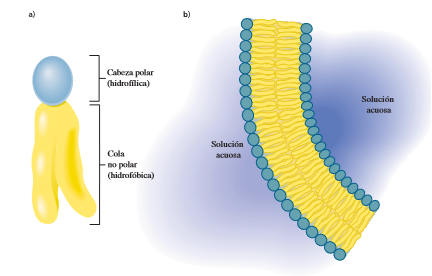 Las bicapas lipídicas son el principal componente estructural de todas las membranas biológicas, incluyendo membranas plasmáticas y membranas internas de células eucariotas. Las interacciones no covalentes entre las moléculas de lípidos en las bicapas hacen que las membranas sean flexibles y les permiten autosellarse. Los triacilgliceroles, que son muy hidrofóbicos y no son anfipáticos, no pueden formar bicapas, y el colesterol, aunque es ligeramente anfipático, no forma bicapas por sí mismo. Una bicapa lipídica tiene en forma típica de 5 a 6 nm de espesor, y consiste en dos hojas o monocapas (que también se llaman hojillas). En cada hoja, los grupos polares de cabeza en los lípidos anfipáticos están en contacto con el medio acuoso, y las colas no polares de hidrocarburos apuntan hacia el interior de la bicapa. La formación espontánea de bicapas lipídicas es impulsada por las interacciones hidrofóbicas (sección 2.5D). Cuando se asocian las moléculas de lípido, la entropía de las moléculas de solvente aumenta y eso favorece la formación de la bicapa lipídica.
Las bicapas lipídicas son el principal componente estructural de todas las membranas biológicas, incluyendo membranas plasmáticas y membranas internas de células eucariotas. Las interacciones no covalentes entre las moléculas de lípidos en las bicapas hacen que las membranas sean flexibles y les permiten autosellarse. Los triacilgliceroles, que son muy hidrofóbicos y no son anfipáticos, no pueden formar bicapas, y el colesterol, aunque es ligeramente anfipático, no forma bicapas por sí mismo. Una bicapa lipídica tiene en forma típica de 5 a 6 nm de espesor, y consiste en dos hojas o monocapas (que también se llaman hojillas). En cada hoja, los grupos polares de cabeza en los lípidos anfipáticos están en contacto con el medio acuoso, y las colas no polares de hidrocarburos apuntan hacia el interior de la bicapa. La formación espontánea de bicapas lipídicas es impulsada por las interacciones hidrofóbicas (sección 2.5D). Cuando se asocian las moléculas de lípido, la entropía de las moléculas de solvente aumenta y eso favorece la formación de la bicapa lipídica.Modelo fluido de mosaico para membranas biológica
Una membrana biológica típica contiene de un 25 a un 50% de lípidos, y de un 50 a un 75% de proteínas, en masa, con menos de 10% de carbohidratos como componente de glicolípidos y glicoproteínas. Los lípidos son una mezcla compleja de fosfolípidos, glicoesfingolípidos (en animales) y colesterol (en algunos eucariotas). El colesterol y algunos otros lípidos que por sí no forman bicapas (30% del total) están estabilizados en el arreglo de bicapa por el otro 70% de los lípidos en la membrana. Las composiciones de las membranas biológicas varían en forma considerable entre las especies, y aun entre distintos tipos celulares en organismos multicelulares. Por ejemplo, la membrana de mielina que aísla a las fibras nerviosas contiene relativamente poca proteína. En contraste, la membrana mitocondrial interna es rica en proteínas, lo que refleja su alto nivel de actividad metabólica.

Las bicapas lipídicas y las membranas son estructuras dinámicas
Los lípidos en una bicapa están en movimiento constante, dando a las bicapas lipídicas muchas de las propiedades de los fluidos. Los lípidos tienen varios tipos de movimiento molecular dentro de las bicapas. El rápido movimiento de los lípidos dentro del plano de una monocapa es un ejemplo de difusión lateral bidimensional.
Las propiedades fluidas de las bicapas lipídicas dependen de la flexibilidad de sus cadenas de acilo graso.
Tres clases de proteínas de membrana
 Las proteínas integrales de membrana, llamadas también proteínas transmembranales, contienen regiones hidrofóbicas incrustadas en el interior hidrofóbico de la bicapa lipídica.
Las proteínas integrales de membrana, llamadas también proteínas transmembranales, contienen regiones hidrofóbicas incrustadas en el interior hidrofóbico de la bicapa lipídica.- Las proteínas periféricas de membrana se asocian a una cara de la membrana, mediante interacciones de carga a carga y con puentes de hidrógeno, con las proteínas integrales de membrana o con los grupos de cabeza polar de los lípidos de membrana.
- Las proteínas de membrana ancladas a lípidos están unidas a una membrana mediante un enlace covalente con un ancla lipídico.

Transporte de membrana
Las membranas plasmáticas separan físicamente una célula viva de su ambiente. Además, dentro de las células eucariotas, las membranas rodean a diversos compartimientos, como al núcleo y a las mitocondrias. Las membranas son barreras de permeabilidad selectiva que restringen el paso libre de la mayor parte de las moléculas. Sin embargo, las de agua, oxígeno y otras pequeñas deben poder entrar a todas las células y moverse libremente entre los compartimientos en el interior de las células eucariotas. Las moléculas más grandes, como las de proteínas y ácidos nucleicos, también deben transportarse atravesando las membranas, incluyendo las que hay entre compartimientos. Las moléculas hidrofóbicas y las moléculas pequeñas sin carga pueden difundirse libremente a través de las membranas biológicas, pero el interior hidrofóbico de la bicapa presenta una barrera casi impenetrable frente a la mayor parte de las especies polares o cargadas.
El tráfico de moléculas polares y de iones a través de las membranas es mediado por tres tipos de proteínas integrales de membrana: canales y poros, transportadores pasivos y transportadores activos. Esos sistemas de transporte tienen distintas propiedades cinéticas y necesidades de energía.
- Termodinámica del transporte en la membrana.
- Los porosy los canales son proteínas transmembranales con un paso central para iones y moléculas pequeñas.
- Transporte pasivo. Las proteínas de transporte pasivo y activo se unen en forma específica a solutos, y los transportan a través de las membranas. En el transporte pasivo, el soluto se mueve a favor de su gradiente de concentración, y el potencial de membrana es favorable.



- Transporte activo. El transporte activo se parece al transporte pasivo en el mecanismo y propiedades cinéticas generales. Sin embargo, el transporte activo requiere energía para mover un soluto contra su gradiente de concentración.
 |
| transporte activo secundario en animales |
- La endocitosis es el proceso mediante el cual las macromoléculas son rodeadas por la membrana plasmática, y son llevadas al interior de la célula dentro de una vesícula lipídica. La endocitosis mediada por receptor comienza con la unión de macromoléculas a proteínas receptoras específicas en la membrana plasmática de la célula
- La exocitosis se parece a la endocitosis, pero la dirección del transporte es la inversa. Durante la exocitosis, los materiales destinados a ser secretados de la célula se encierran en vesículas mediante el aparato de Golgi.
No hay comentarios:
Publicar un comentario